
Supporting the community
Supporting the community
Alison Kraigsley si Paul D. Ronney
Departamentul de Inginerie Aerospatiala si Mecanica
Universitatea din California de Sud, Los Angeles, CA 90089-1453
Steven E. Finkel
Departamentul de Stiinte Biologice
Universitatea din California de Sud, Los Angeles, CA 90089-1340
Abstract
Desi se cunosc multe despre fronturile de reactie-difuzie auto-propagatoare care apar in multe sisteme chimice care reactioneaza, cum ar fi flacarile, procesele de polimerizare si unele reactii apoase, aceasta baza vasta de cunostinte nu a fost aplicata sistematic in sistemele biologice cum ar fi raspandirea bacteriilor molate. Scopul acestei lucrari este (1) de a determina daca astfel de cunostinte pot fi aplicate sistemelor biologice si (2) daca este cazul, sa obtina o intelegere mai cantitativa si predictiva a sistemelor biologice care prezinta fronturi auto-propagatoare. Analogia dintre raspandirea bacteriilor molate si a altor fronturi auto-propagatoare va fi urmarita cu privire la proprietatile dinamice ale unor astfel de fronturi. Experimentele initiale care utilizeaza bacteria E. coli arata intr-adevar un comportament analog cu sistemele de difuzie de reactie. Incercam sa realizam un studiu cuprinzator al proprietatilor dinamice, incluzand ratele de propagare, efectele curburii frontale, limitele de stingere, limitele de stabilitate, efectele de flotabilitate, coeficientii de difuzie efectivi si scala timpului de reproducere. Aceste informatii vor fi folosite ca intrari pentru modelele analitic / numerice existente ale fronturilor de difuzie a reactiilor si validitatea analogiilor propuse determinate. Diferentele dintre sistemele conventionale de difuzie de reactie si sistemele microbiologice vor fi, de asemenea, evaluate. O diferenta esentiala este ca bacteriile evolueaza si raspund la stres, un factor care va fi studiat prin repetarea testelor de mai sus cu bacterii care au supravietuit conditiilor de stingere aproape. Aceste date vor fi apoi utilizate pentru a obtine o mai buna intelegere a supravietuirii si adaptarii bacteriilor in conditii adverse.
Introducere
Frunzele de reactie auto-propagatoare apar in multe sisteme chimice si fizice, incluzand flacari, procese de polimerizare initiate de radicali liberi si unele reactii apoase. Toate aceste sisteme se caracterizeaza prin doua caracteristici principale: un mediu reactiv (de exemplu un amestec combustibil-aer in cazul flacarilor) si un autocatalizator care este un produs al reactiei care accelereaza de asemenea reactia (de exemplu, energia termica in caz de flacari). Auto-propagarea apare atunci cand autocatalizatorul difuzeaza in mediul reactiv, initiand reactia si creand mai mult autocatalizator. Acest lucru permite ca fronturile de difuzie a reactiei sa se propageze la rate stabile, departe de orice loc de initiere.
Doua dintre caracteristicile cheie ale fronturilor de reactie-difuzie auto-propagatoare sunt viteza de propagare si limita de stingere. Rata de propagare este viteza cu care frontul avanseaza in mediul reactiv, care in general depinde de scala globala a timpului de reactie (t) si de coeficientul de difuzie (D) al autocatalizatorului in functie de relatia s ≈ (D / t) 1 / 2. T in functie depinde de concentratia si activitatea reactantilor. Limita de stingere este valoarea minima sau maxima a unui anumit parametru (de exemplu, concentratia minima a reactantilor sau latimea canalului) pentru care poate exista o fata propagatoare constanta. Cuplajul care are loc intre reactiile chimice si transportul difuziv si convectiv in flacari [[1]], fronturile de polimerizare [[2]] si reactiile apoase [[3]] este bine inteleasa cantitativ.
In mod surprinzator, linia analogica de ancheta nu a fost aplicata sistemelor microbiologice, desi prima modelare a fronturilor difuzive reactive, care conduce la relatia s ≈ (D / t) 1/2, a fost realizata in 1937 in contextul raspandirii bolilor infectioase (vezi [[4]]). Scopul cercetarii noastre este de a evalua fezabilitatea efectuarii acesteia si de a dezvolta o intelegere mai cantitativa a unor astfel de sisteme microbiologice. Majoritatea studiilor microbiologice se concentreaza pe bacteriile individuale si pe comportamentul indivizilor. In acest studiu se aplica legi termodinamice si se studiaza comportamentul agregat al unui numar mare de indivizi. Cazul de testare pe care alegem sa-l concentram asupra acestui studiu este bacteria Escherichia coli foarte frecventa si studiata, o bacterie motila care (ca multe altele) inoata folosind tentaculele sau flagelul in mediile sale nutritive, schimband in mod frecvent directia pentru a cauta regiuni mai inalte Concentratia nutrientilor. In mod specific, bacteria are doua moduri de comportament: modul "alergare" in care rola sa se roteste pentru a o propulsa intr-o linie mai mult sau mai putin dreapta si modul "tambur" in cazul in care flagella provoaca schimbarea orientarii cu putina miscare neta [5] . Miscarea rezultata este oarecum analoga cu plimbarea aleatorie a moleculelor care conduce la difuzia clasica Fickian (desi mecanismul este complet diferit, deoarece, spre deosebire de molecule, miscarea bacteriilor nu este legata de coliziuni elastice intre bacterii). De asemenea, intr-un gradient favorabil al nutrientilor, bacteria ruleaza mai mult decat se misca, rezultand in D mai mare, in timp ce in gradiente nefavorabile, se rupe mai mult, permitandu-i sa caute noi directii si ducand la scaderea D [5]; Acest lucru conduce, de asemenea, la o plimbare aleatorie partinitoare care favorizeaza migrarea in regiuni cu niveluri ridicate de nutrienti.
Aceste caracteristici ale bacteriilor molate pot fi exploatate pentru a modela comportamentul lor ca fronturi de reactie-difuzie. Viteza de propagare a E. coli masurata (vezi sectiunea rezultate preliminare) intr-un mediu de agar de 0,3% este de aproximativ 4,5 mm / ora. Scala timpului de reproducere (t) a E. coli este de aproximativ 20 min. 5. Deoarece viteza de propagare s ≈ (D / t) 1/2, D ≈ s2t, deci D ≈ 1,5 x 10-5 cm2 / sec. Aceasta valoare este in concordanta cu valoarea asteptata pe baza teoriei cinetice a moleculelor care arata ca D este proportionala cu viteza particulelor (c) inmultita cu calea libera a particulelor (l), adica in cazul moleculelor distanta dintre particule Inainte de a se ciocni unul cu celalalt sau, in cazul bacteriilor motila, schimbarea directiei. Poate fi estimat ca c inmultita cu timpul (t) de inot al bacteriile fara a schimba directia. Pentru E. coli, a fost masurata o viteza medie de inot de 21 μm / s, cu 21 modificari in directie pe 30 de secunde [5]. Aceasta implica ≈ 1.4 s si l ≈ 3.0 x 10-3 cm si, prin urmare, difuzia efectiva D ≈ 6.3 x 10-6 cm2 / s, care are aceeasi ordine de marime ca si valoarea D dedusa din D ≈ s2t .
Concluzionam ca este rezonabila caracterizarea raspandirii bacteriilor molate ca fronturi de reactie-difuzie. Multe studii microbiologice masoara raspunsul sistemului la un gradient de temperatura sau nutrienti care este impus sistemului; Intentionam sa aratam ca bacteriile isi pot genera propriile gradienti si, prin urmare, pot fi auto-propagatoare. Tabelul 1 prezinta analogia propusa intre flacari (o fata tipica a difuziei de reactie) si sisteme microbiologice.
Flacara sau proprietati moleculare | Echivalent echivalent microbiologic |
| Temperatura | Concentratia bacteriilor |
| Combustibil | Nutrienti |
| Difuzivitatea caldurii≈ cl | Difuzivitatea bacteriilor |
| Difuzivitatea combustibilului | Difuzivitatea nutrientilor |
| Viteza sunatului (c) | Viteza de inot a bacteriei in modul "alergat" |
| Calea libera medie (l) | c multiplicat cu timpul mediu pentru trecere de la mediul de functionare la modul tambur si inapoi |
| Timpul de reactie (t) | Timpul de reproductie |
| Pierderea de caldura | Moartea (a unei bacterii) |
| Stoparea | Moartea (a tuturor bacteriilor) |
Tabelul 1. Analogie propusa intre flacari si fronturi microbiologice | |
Un factor complicat in flacari, precum si bacteriile molate este acela ca coeficientul de difuzie eficient se modifica odata cu concentratia produsului. In cazul flacarilor, productia de produse duce la o temperatura mai ridicata care creste D in aproape toate cazurile. In cazul bacteriilor motile, este bine cunoscut faptul ca bacteriile petrec mai putin timp in modul "alerga" si mai mult in modul "tambur" atunci cand concentratia de nutrienti este ridicata (de ce priviti cand iarba este deja verde?) Inseamna ca traiectoria medie libera va fi mai mica si astfel difuzia eficienta va fi mai mica atunci cand concentratia nutrientului este ridicata. In cazul unei fete de inmultire, cele cateva bacterii de pe muchia frontala a frontului vad concentratii mari de nutrienti si, prin urmare, au un nivel scazut de D. Concentratiile mai mari de bacterii ("produsul" reactiei) la marginea posterioara a frontalului vor exista in Un mediu cu o concentratie mai scazuta de nutrienti si astfel va "rula" mai mult si mai putin "tambur", crescand l si astfel D va creste. Astfel, analogia cu flacarile este destul de larga.
Discutia de mai sus ne-a incurajat sa realizam experimente preliminare pentru a testa viabilitatea propunerilor noastre. Aceste experimente s-au efectuat la 37 ° C in placi standard Petri utilizand un mediu nutritiv pe baza de apa constand din 1% NaCI, 1% tryptona, 0,5% drojdie si 0,1% agar (daca nu se mentioneaza altfel). Agarul mareste viscozitatea mediului si, in esenta, previne orice miscare in vrac. Mediul a fost inoculat cu bacterii intr-un punct (de obicei in centrul vasului), iar avansul (daca exista) al fetei bacteriene a fost observat vizual. Au fost testate mai multe tulpini de E. coli. Dupa o perioada tranzitorie initiala, toti au prezentat o crestere liniara a razei frontale cu timpul. Un exemplu este prezentat in Fig. 1.
Figura 1 arata ca frontul avanseaza mai incet initial atunci cand raza din fata este mica si curbura frontala inalta. Toate celelalte teste au aratat aceeasi tendinta. Acest comportament este observat pe fronturile chimice in care produsul autocatalitic are o difuzivitate mai scazuta decat cea a reactantului (adica, in cazul flacarilor, o difuzivitate termica ridicata si o difuzivitate scazuta a combustibilului) si motivele sunt bine intelese [6] ]. Desi difuzivitatea nutrientului de triptona nu este cunoscuta, prin analogie cu molecule similare poate fi estimata o valoare de 10-7 cm2 / s, care este mai mica decat difuzia aparenta a E. coli asa cum s-a estimat mai sus. In consecinta, comportamentul prezentat in Fig. 1 este in concordanta cu modelul de difuzie a reactiei de propagare frontala. Mai mult decat atat, teoria flacarii [1] arata ca, daca raportul difuzivitatii produsului la difuzitivitatea reactantului este prea diferit de valoarea 1, fie mai mare, fie mai mica, pot rezulta diferite tipuri de instabilitati frontale. Formarea formelor in sistemele microbiologice este bine cunoscuta [[7]], dar teoria difuziei de reactie nu a fost utilizata pentru a cuantifica si prezice astfel de modele. Vom investiga astfel de instabilitati prin modificarea difuziei medii prin concentratia de agar. Mai mult decat atat, suspectam ca aceasta alegere a timpului de rulare a E. coli nu este accidentala deoarece afecteaza D; Daca D este prea mic sau prea mare, rezulta instabilitati care pot fi nefavorabile pentru supravietuire. Vom urmari aceasta notiune examinand efectul concentratiei de agar asupra vitezei si timpului de rulare a bacteriilor sub microscop si determinand daca bacteria se adapteaza (fie in zbor, fie prin procese evolutive), viteza si timpul de rulare Pentru a modifica D efectiv pentru a mentine fronturi stabile.
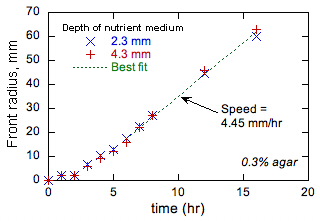 |
Figura 1. Exemple de date experimentale privind razele frontale bacteriene in functie de timp. |
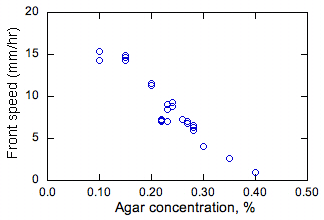 |
Figura 2. Efectul masurat al agarului si adancimii solutiei asupra vitezei de propagare constanta a fronturilor bacteriene. |
Efectul concentratiei de agar si adancimea mediului nutritiv asupra valorilor constante ale vitezei de propagare s este prezentat in Fig. 2. La o concentratie de agar de 0,5% sau mai mare (nu este prezentata), mediul a fost in esenta solid si bacteriile se raspandesc numai pe suprafata mediului, nu in vrac. Diferitele comportamente ale bacteriilor sunt putin probabile in acest caz, deoarece nu pot inota. La concentratii de agar de 0,4% si mai scazute, bacteriile se raspandesc prin mediu. Cele mai mari rate de propagare au fost observate la cele mai scazute concentratii de agar (si astfel, cea mai mica vascozitate a mediului). Acest lucru este de asteptat deoarece, la o vascozitate mai scazuta, bacteria poate inota mai repede si astfel ar trebui sa aiba D mai mare, ducand la s mai mare (deoarece s este proportionala cu D1 / 2 asa cum s-a discutat mai sus).
S-a constatat ca adancimea solutiei nu are aproape niciun efect asupra s. Prin realizarea sectiunilor transversale ale mediei dupa propagarea partiala a frontului, s-a constatat ca fronturile sunt in mare parte uniforme in dimensiunea verticala. Astfel, fluxul de flotabilitate nu afecteaza aceste fronturi, altfel ar fi de asteptat un efect al adanciturii solutiei asupra s sau al unei neuniformitati in directia verticala. Deoarece testele au fost efectuate in aer, aceste observatii indica, de asemenea, ca difuzia de oxigen in mediu nu este semnificativa, in caz contrar cresterea in apropierea suprafetei solutiei nutritive ar fi diferita de cea care se afla sub suprafata. Acest ultim rezultat arata ca bacteriile cresc anaerob.
Pentru a determina daca limitele "quenching" exista in fronturile bacteriene, a fost efectuat un experiment in care fronturile bacteriene au incercat sa se propageze prin canale inguste si largi cu pereti antibiotici. Deoarece antibioticele elimina produsul autocatalitic (bacteriile), efectul peretilor este similar cu pierderea de caldura la peretii reci in flacari. Pentru propagarea frontala in canalele inguste, este bine cunoscut [1] ca apare o limita de caldura datorita pierderilor de caldura la peretii canalului atunci cand numarul Peclet Pe = sw / D, unde w este latimea canalului si D este difuzia, este Mai putin decat o valoare critica. Astfel, in prezenta pierderilor de caldura, fronturile se pot propaga prin canale largi (pe inaltime Pe), dar nu pe canale inguste (joase Pe). Figura 3 arata ca fronturile se pot propaga prin canalul larg, dar nu pe canalul ingust, indicand o limita de stingere. Aplicabilitatea relatiei de stingere Pe = sW / D = constanta la limita va fi testata in aceasta lucrare.
A fost de asemenea testat efectul concentratiei nutrientului de triptona; s-a constatat ca la jumatate din valoarea nominala nu s-ar produce nici o raspandire, in timp ce s a fost aproape neschimbat prin dublarea concentratiei de triptona fata de valoarea nominala. Acest lucru poate indica o limita de stingere chiar si in absenta zidurilor cu antibiotice. Un comportament similar apare in flacari datorita pierderilor de caldura prin radiatie atunci cand concentratia de combustibil este prea mica. Vom investiga aceasta limita fara perete si vom incerca sa determinam mecanismul de pierdere responsabil pentru suprimarea propagarii frontale la niveluri scazute de nutrienti.
Titrarea mediului pentru a obtine numarul de bacterii a aratat in mod constant un maxim semnificativ in concentratia bacteriilor de pe frontul principal, urmata de o scadere importanta in spatele frontului si, pentru perioade ulterioare, de un al doilea varf, cu mult inaintea primului. Acest lucru este in concordanta cu imaginile prezentate in Fig. 3. In lucrarile viitoare vom folosi analiza de titrare pentru a determina daca grosimea frontala ≈ D / s este prezis teoretic [1]. Pe baza examinarii vizuale din Fig. 3 grosimea frontala este de aproximativ 1 mm, care este aproape de predictie D / s = (1,5 x 10-3 mm2 / sec) / (4,5 mm / h) = 1,2 mm. Rezultatele cu o tulpina salbatica de bacterii au prezentat o tranzitie initiala putin mai rapida si un timp si o distanta mai scurta pentru a atinge o rata constanta de propagare in comparatie cu tulpinile marcate genetic.

Figura 3. Fotografiile fronturilor bacteriilor motile E. coli propagate prin canale inguste (6 mm, stanga) si late (35 mm, dreapta) cu pereti laterali antibacterieni kanamycin (100 microliti Kanamycin pe fiecare parte), luati la 6,5 ore dupa inoculare. Ambele cazuri: 2086 tulpina salbatica de E. coli, 0,1% agar, mediu nutritiv standard .
Program actual de cercetare
Pe baza succesului aparent al modelului de difuzie de reactie in descrierea in aceste rezultate experimentale preliminare, urmarim urmatorul program de cercetare:
Desigur, exista, de asemenea, unele diferente intre sistemele conventionale de difuzie de reactie si sistemele microbiologice. O diferenta esentiala este ca bacteriile evolueaza si raspund la stres, un factor care va fi studiat prin repetarea testelor de mai sus cu bacterii "experimentate" care au supravietuit conditiilor de stingere aproape. O alta diferenta, asa cum s-a discutat mai sus, este proprietatile de difuzie partinitoare. Aceasta prejudecata poate fi modelata folosind o relatie de difuzie neliniara pentru a inlocui Legea lui Fick.
Importanta practica
Aceasta lucrare este un studiu al dinamicii unei noi clase de fronturi propagatoare care implica bacterii motile. Studiul dinamicii din fata are multe aplicatii, dupa cum s-a discutat mai sus. Aceste studii, cu toate acestea, prezinta un interes deosebit pentru microbiologi, deoarece se refera la mecanismele de supravietuire si evolutie pe termen lung folosite de bacterii. Unul dintre factorii primari care influenteaza ratele de schimbare evolutiva in aceste sisteme bacteriene sunt nivelurile stresului de mediu, precum si trecerea de la un mediu la altul. Sistemele de cultura pe care le dezvoltam reprezinta medii noi in care nu au fost abordate aceste probleme de evolutie si de supravietuire. Aceasta lucrare ofera prima incercare, spre cunostinta noastra, de a caracteriza aceste efecte biofizic.
Studiul de crestere a E. coli in conditii anaerobe prezinta un interes deosebit. Atunci cand E. coli (si toate celelalte bacterii enterice) cresc in asociere cu tractul digestiv uman, ele cresc in conditii anaerobe. Dorim sa incepem sa studiem raspunsurile stresului bacterian in absenta oxigenului pentru a ne extinde intelegerea generala a modului in care aceasta bacterie se adapteaza la viata intr-o mare varietate de nise de mediu. De exemplu, nu se stie in prezent daca mecanismele repararii si mutatiei daunelor ADN pe care Prof. Finkel le studiaza (si pe care le-a dovedit a fi importante pentru adaptarea la medii noi) functioneaza in conditii anaerobe. Aceste studii vor reduce decalajul de la cercetarea de baza in mai multe domenii relevante din punct de vedere medical.
We believe in helping people and that matter to us more than anything else. Since the very beginning of our company, our team have been willing and wishing to help.